
Evolution and development of embryogenesis
All creatures living in the earth have evolved through modification of their embryonic development. How have they achieved this? I study it.
私は胚発生がどのように進行し、また生物の進化の過程でどのように変化を遂げたかという点に興味を抱き、研究を進めています。これらはそれぞれ「発生生物学 developmental biology」、「進化生物学 evolutionary biology」という元々は異なる研究分野であったものが分子生物学の興隆により融合し、近年では「進化発生生物学 evolutionary developmental biology (evo-devo)」と呼ばれています。私は生物の胚発生とその進化の関係性に迫るために、遺伝子/ゲノム、一細胞レベルでのふるまい、組織構成、器官形成、そして個体としての恒常性と、様々な階層から生命現象にアプローチしています。以下に現在取り組んでいるテーマを紹介します。
Evolution and development of the heart

心臓は胚発生の最も初期から機能し、個体が死に至るまで血液を循環させ続けるという、まさに生命そのものと言ってよい重要な器官です。そのため、全ての生物は生息する環境に適応するように心臓の形態や機能を変化させており、その多様性は生物進化の巧妙さをよく表しています(図1)。では、このような心臓形態・機能の多様性はいったいどのようなメカニズムによって生み出されているのでしょうか?私は胚発生における心臓形態形成とその機能の制御に様々な階層からアプローチし、また異なる生物種の心臓発生様式を相互に比較することにより、心臓の発生と進化に迫っています。

最近の研究結果から
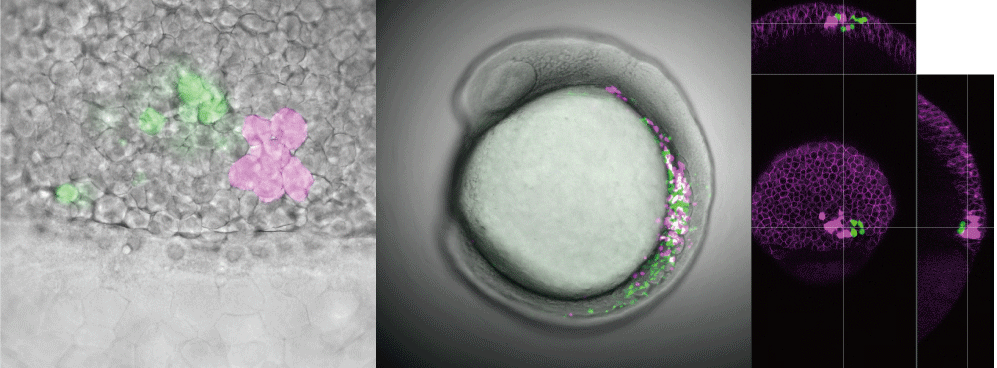
脊椎動物の心臓は大きく4つの部屋から構成されています。血液が流入する側から、静脈洞、心房、心室、流出路です。四肢動物ではこの中の心房、心室をそれぞれ中隔によって隔てることで動脈血と静脈血を分け、循環器系をより洗練されたものへと変化させました(図1)。一方で、水中環境に生息する真骨魚類は別の方法で心臓形態を変化させ、循環器系を洗練させています。真骨魚類を除く脊椎動物では流出路は"動脈円錐"と呼ばれ、心房、心室同様に心筋から構成されているのですが、真骨魚類では流出路が平滑筋から構成されており、"動脈球"と呼ばれる構造となっています(図2)。動脈球はエラスチン繊維に富む弾力性の高い器官で、心室から押し出されてきた血液を一旦受け止め、適当な血圧・血流量に調節して血液を全身に運ぶという機能を担っており、このことから真骨魚類の循環器系は水中環境に生息する魚類の中でも最も効率の良い系となっています。以上のような特徴から、動脈球は真骨魚類の進化における最も重要な進化的新規形質であると考えられていますが、どのような発生学的メカニズムによって形成され、また進化の過程で獲得されたのかはこれまでほとんど明らかにされてきませんでした。

近年、私はelastin bという細胞外マトリックスをコードする遺伝子が真骨魚類の進化の過程で獲得され、この遺伝子の働きによって心臓流出路を構成する細胞が心筋から平滑筋へと細胞運命を変化することで動脈球が形成されることを明らかにしました。真骨魚類では系統特異的な全ゲノム重複(Teleost Specific Whole-Genome Duplication: TSGD, 3rd Round Whole-Genome Duplication: 3R WGDとして知られています)が生じていることが知られていますが、今回発見したelastin bもこの全ゲノム重複で生じたことを明らかにしました。ゲノム重複や遺伝子重複が進化の原動力になるかについては議論の的でしたが、本研究はゲノム重複が確かに進化の原動力となることを示しました。また、elastinは細胞外マトリックスをコードする遺伝子です。このことは、流出路を構成する細胞が、細胞外の環境(弾性や粘弾性など)を感知して自身の細胞運命を変化させていることを示唆しています。実際、メカノトランスデューサーとして知られるYAPの機能阻害をおこなうと、elastin bの機能阻害同様、平滑筋で構成されるはずの流出路構成細胞が心筋へと変化してしまいます。このことからも、上記のような細胞外環境の変化による細胞運命の変化が支持されます。
本研究は生物が進化の過程でどのように新たな器官を獲得し、環境に適応するように胚発生プロセスを変化させたかという、生物学の根源的な問いに迫るものであると同時に、細胞運命をいかに人為的に変化させ、制御するかという医療応用にも繋がるものでもあります。スイス生まれの動物学者、ルイ・アガシ−(Jean Louis Rodolphe Agassiz)の言葉に"Study nature, not books"というものがありますが、私もまた、胚発生の観察とその種間比較から、生命の本質に迫りたいと考えています。
Cell behavior, mechanics and the evolution of gastrulation

全ての左右相称動物はその発生初期に原腸形成を経ることにより、外胚葉、内胚葉、中胚葉といった胚葉を成立させ、体制を作り上げていきます。この原腸形成の様式は進化の過程で著しく変化しており、各系統によって異なる様式をとります(図3)。またその一方で、全ての種の原腸形成は抽象化するとその機構は高度に保存されているとも言えます。つまり、それぞれ外から中へと細胞が移動し、中へ入り込んだ細胞群が内側(卵黄側)から順に内胚葉、中胚葉となり、外側に残った細胞群が外胚葉となり、原腸形成完了時には3胚葉が成立しているという点で、全ての原腸形成は様式を変えながらもその本質は高度に保存されているとも言えます。私はゼブラフィッシュをモデルとして、原腸形成における細胞の挙動やその物性的性質を、ライブイメージングを駆使することによって明らかにしていこうとしています。また、ここで明らかとなった分子メカニズムについて、他の生物種と比較することで、原腸形成のどのプロセスが進化の過程で変化しうるのか、つまり"原腸形成期における発生拘束(developmental constraint for the evolution of gastrulation)"について明らかにしていきたいと考えています。

Gene duplication as a means of morphological/functional evolution

Evolution by Gene Duplication(1970)において、遺伝子重複が進化の原動力であるという仮説を提唱し、現在までにそれを支持する研究成果が報告されています。では、一体どのような遺伝子重複が、どのような形態や機能の進化をもたらしているのでしょうか?重複した遺伝子の多くは変異の蓄積により偽遺伝子(pseudogene)となってしまうため、この点についてはいまだ十分な理解は得られていません。
上記に挙げた脊椎動物の心臓進化に関する研究の過程で、私はelastin遺伝子の重複とその姉妹遺伝子の新規機能獲得(neo-functionalization)が真骨魚類に動脈球をもたらしたことを明らかにしました。また、elastin遺伝子は真骨魚類特異的な全ゲノム重複(3R WGD)によって重複していたことから、3R WGDが実際に進化の原動力になっているということも示されました。私はこの3R WGDに着目し、どのような遺伝子の重複が真骨魚類の特徴的な形質の進化に寄与しているのかを研究しています。
Acquisition of dorsal identity and the evolution of caudal skeleton

上記の心臓の例と同様に、体を構成する様々なパーツもまた、生息する環境に適応するようにその形態を変化させています。真骨魚類は水中環境に適応することで爆発的に種数を増やし、繁栄を遂げました。遊泳能力を向上させ、水中の様々なニッチに適応することができたのは脊柱後端部が背側に屈曲することで形成される"正型尾"の獲得であると考えられています。正型尾は真骨魚類に特有の形態であり、水中環境での遊泳に最も適した形態であると考えられています。外見上は背腹対称ですが、内部構造は極端に背腹軸に対して非対称な形態となっています。骨格としては、尾部骨と呼ばれる脊椎後端部が背側に屈曲し、下尾骨と呼ばれる特殊化した腹側の骨要素が尾鰭を支える形態となっています。以上のように、正型尾は真骨魚類の進化における重要な進化的新規形質の一つと考えられていますが、どのようなメカニズムによって形成されるかについてはほとんど明らかとなっていませんでした。私は、体幹部・尾部の背腹非対称性が失われる表現型を示すメダカ自然突然変異体Double anal fin (Da)や、正型尾をもたない条鰭類に着目することで、正型尾の形成メカニズムに迫っています。
これまでの研究結果から
Daの解析から、Daは転写因子zic1/zic4の体幹部、尾部のエンハンサー活性が巨大なトランスポゾンの挿入により失われているエンハンサー変異体であり、そのために体幹部、尾部の形態でのみ背側のアイデンティティが消失していることを明らかにしました。また、真骨魚類胚では一過的に体軸後端部まで体節が形成されるものの、その後後方の体節が融合し、それによって硬節に富む、体幹部とは異なった特殊な組織構造(caudal-skeleton forming mesenchyme: CSM)が形成されることを見出しました。以上から、脊椎動物の進化の過程において、真骨魚類に至る系統においてCSMが獲得され、またzic1/zic4が背側CSMへco-optionされることにより背腹非対称な尾部骨格とそれによる正型尾が獲得されたという進化シナリオが示唆されました。
Morphological evolution of paired appendages
四肢動物のもつ手足と、魚類がもつ対鰭(胸鰭、腹鰭)はそれぞれ相同器官であり、共通の起源を有しています。脊椎動物の進化の過程では、水棲であったものがある時点で陸に上がり、四肢を獲得したと考えられています。この際の鰭から四肢への変化はfin to limb transitionと呼ばれ、現在盛んに研究が進められている分野です。私は四肢と鰭の発生プロセスの共通性、相違性を探ることによりその進化プロセスに迫っています。
References:
Moriyama et al., Zoological Science. 2019
Kawahata et al., Scientific Reports. 2019
Okamoto and Moriyama et al., Development, Growth and Differentiation. 2019